“啤酒酵母”的版本间的差异
| (未显示同一用户的16个中间版本) | |||
| 第176行: | 第176行: | ||
:*细胞膜 | :*细胞膜 | ||
::紧贴细胞壁的内面,厚度约150nm,是一层半透性的膜,构成细胞壁的基础物质。细胞膜调节着细胞内的渗透压,调节着营养物质的吸收和代谢物的排出,形成酵母细胞的渗透框架。同时,细胞膜可分离出[http://baike.baidu.com/link?url=zPr0jJlEa8uqOhftTItwqp-fO39DcuwT_u4v-k0eMu0SL7K3jIaUMjGYDsWhDFPZ36kzQfNyM1G4wvRKfl2r6gciQJz3HnNCmx8U3EHReWZ6XpcvH4n98zf8tUQSE8L- 胞外酶],胞外酶由酵母细胞形成,但在酵母细胞外起作用。 | ::紧贴细胞壁的内面,厚度约150nm,是一层半透性的膜,构成细胞壁的基础物质。细胞膜调节着细胞内的渗透压,调节着营养物质的吸收和代谢物的排出,形成酵母细胞的渗透框架。同时,细胞膜可分离出[http://baike.baidu.com/link?url=zPr0jJlEa8uqOhftTItwqp-fO39DcuwT_u4v-k0eMu0SL7K3jIaUMjGYDsWhDFPZ36kzQfNyM1G4wvRKfl2r6gciQJz3HnNCmx8U3EHReWZ6XpcvH4n98zf8tUQSE8L- 胞外酶],胞外酶由酵母细胞形成,但在酵母细胞外起作用。 | ||
| + | ::细胞膜中最重要的基础物质是磷脂,磷脂具有对其他作用而言非常重要的典型结构。每两个脂肪酸残基与甘油产生酯化反应,在甘油的第三个OH<sup>-</sup>根上,通过磷基结合了一个氨基酸(磷脂) | ||
| + | ::脂肪是细胞膜的主要组成物质,其形成过程消耗很多能量,同时需要氧气存在,在此过程中脂肪酸的一部分转化为熔点较低、运动性更好的不饱和脂肪酸,缺乏氧气时细胞的形成会提前停止。 | ||
:*细胞质 | :*细胞质 | ||
| − | :: | + | ::占细胞体积50%以上,是细胞内部最重要的部分,是细胞的反应中心,大多数营养物质的分解代谢及细胞自身基础物质的合成过程都在这里完成,整个中间物质代谢:糖解、脂肪酸的分解、蛋白质的生物合成以及其他过程都在这里同时进行。 |
| + | ::营养物质丰富的时候,如发酵开始时,酵母细胞的储存物质会增加,糖原这种储备碳水化合物的含量可超过酵母干物质的30%,储存在细胞质中,海藻糖这种双糖也会储备,此外还有酵母形成新的细胞物质所必需的磷和脂肪。 | ||
| + | ::细胞中通常会充满酸性细胞液和液泡,这里储存着特定的蛋白质和多余的盐类,部分为结晶形式,当外部的渗透压由于较高的浸出物或酒精含量上升时,细胞可以通过结晶盐的可逆変化调整其内部压力。 | ||
| + | |||
:*细胞核 | :*细胞核 | ||
| 第210行: | 第215行: | ||
:两个酵母菌通过接触并融合后形成子囊孢子,子囊孢子在十一的环境下萌发,然后孢子分离母细胞后开始生长,并重新以出芽的方式进行繁殖。 | :两个酵母菌通过接触并融合后形成子囊孢子,子囊孢子在十一的环境下萌发,然后孢子分离母细胞后开始生长,并重新以出芽的方式进行繁殖。 | ||
:[[文件:酵母的有性繁殖.jpg]] | :[[文件:酵母的有性繁殖.jpg]] | ||
| + | |||
| + | *酵母的生长 | ||
| + | :酵母菌在生长繁殖过程中也遵循幼而壮,壮而老,老而衰,衰而死的生物生长规律。把一定数量的酵母菌,接种于合适的液体培养基中,在适宜的温度下培养时,它的生长过程具有一定的规律性。在其生长过程中,定时取样计算酵母细胞数量,然后以酵母细胞数的对数作纵坐标,生长时间作横坐标,绘制所得的曲线图称为生长曲线。 | ||
| + | :[[文件:酵母菌的生长曲线.jpg]] | ||
| + | |||
| + | :分析酵母的生长曲线,大致可划分为四个阶段: | ||
| + | ::*迟缓期 | ||
| + | :::酵母菌刚接入新的培养液中,一般并不立即开始生长,需要经过一定的时间才开始生长,这是由于细胞内各种酶系要有一个适应的过程,这段时间就叫做迟缓期或者调整期。迟缓期的长短与接入菌种的老幼、培养液的成分、菌种的种类有关。如果菌种来自生长旺盛的培养液,麦芽汁温度适宜,适当充气供氧,麦汁营养充足,则迟缓期短,反之则长。在此期间,酵母细胞并不增加(即增值速度为零),但每个细胞体积增大,细胞质变得更加均匀,贮藏颗粒物质逐渐消失,代谢机能变得非常活跃,然后开始出芽繁殖,生长速度逐渐加快,时间可能需要数小时。 | ||
| + | :::在生产中,为缩短生长周期,通常希望调整期时间短一些,可以通过增加接种数量、采用生理态处于对数生长期的酵母细胞做种子等措施来实现。 | ||
| + | |||
| + | :::''这里可以添加一个“加速期”,此阶段紧接着迟缓期,细胞分离速度加快'' | ||
| + | |||
| + | ::*对数生长期 | ||
| + | :::酵母经过迟缓期后,就以最快的速度进行繁殖,进入对数生长期。此时,酵母每增殖一次,所需时间最短,酵母在恒定时间内,每个细胞进行芽殖,由一个细胞变两个,两个细胞变四个,即以2<sup>n</sup>的速度增长,指数n代表酵母细胞增殖世代(细胞生殖的一个周期)的数目。无论开始时细胞数目多少,其繁殖率总是2<sup>n</sup>。新细胞增殖所需要的时间称为世代时间(G),繁殖的每个世代的时间随菌种和培养条件相差甚大。 | ||
| + | :::如果酵母培养的总时间为t,以Xo表示接入培养基的酵母数,经过t时间培养后,测得其细胞总数为Xt。公式:Xt = Xo × 2<sup>n</sup> | ||
| + | ::::由此可求出n,即在t时间内增殖世代数: | ||
| + | :::::lgXt = lgXo + nlg2 | ||
| + | :::::n = (lgXt - lgXo) / lg2 | ||
| + | ::::: = 3.32lg(lgXt / lgXo) | ||
| + | ::::因为世代时间 G = t / n,由此可求: | ||
| + | :::::G = t / (3.32lg(Xt / Xo)) | ||
| + | :::在对数生长期中,酵母细胞数目的增加与时间成正比关系。在对数生长阶段,世代时间是稳定的。酵母在最适条件下生长的世代时间为2小时。影响世代时间的主要因素除菌种本身外,营养物质的浓度是主要的,此外还有培养温度、ph、渗透压和代谢产物浓度等因素。处于对数生长期的酵母细胞,其个体形态和生理特性比较正常,代谢旺盛,生长速度恒定。 | ||
| + | |||
| + | :::''这里可以添加一个“稳定期”,此阶段紧接着对数生长期,由于各种因素,比如底物减少,抑制生长的代谢物增加等,此阶段酵母细胞的增值速度逐渐减小'' | ||
| + | |||
| + | ::*稳定期 | ||
| + | :::酵母菌体培养到一定时间大量繁殖后,培养基内的营养物质逐渐耗尽,代谢产物(如酒精、二氧化碳等)积累,ph也改变,使之对细胞快速增值产生抑制作用,细胞不再以指数率无限地生长,自溶数和繁殖数两者达到平衡,维持一个动态平衡,使酵母细胞数维持为恒定值。 | ||
| + | |||
| + | ::*衰亡期 | ||
| + | :::酵母菌生长到后期,培养基中养分大量消耗,代谢产物更多,环境条件对酵母生长逐渐不利。此时酵母细胞大量衰老,细胞自溶不断增多,活酵母逐渐减少。末期,繁殖几乎等于零,死亡率急剧增加。 | ||
| + | |||
| + | :::生产中为了缩短发酵时间,往往采取各种措施使酵母及早进入对数生长期。如在酵母扩培阶段直接扩培至生产规模,选择在酵母增殖的对数生长期进行接种,以缩短酵母增殖的调整期。 | ||
| + | |||
| + | ===酵母所需的营养=== | ||
| + | 酵母细胞由多种化学物质组成。其中最主要的是蛋白质、核酸、多糖和脂类这四类大分子,他们约占细胞干重的96%。其余就是组成它们的单体以及无机盐等。水约占70%,此外还有有机酸、维生素、激素等有机化合物。这些化学物质均由碳、氢、氧、氮、磷、硫以及其他为数不多的化学元素构成。这些化学元素都来自于胞外环境。酵母细胞利用含这些化学元素的物质制造其细胞物质和组分,并进一步将它们组织成细胞结构。 | ||
| + | 在微生物的营养中有以下6大要素物质,它们是:碳源、氮源、能源、生长因子、无机盐、水。 | ||
| + | |||
| + | *碳源 | ||
| + | :碳是微生物细胞需要量最大的元素。能提供微生物营养所需碳元素的营养物质称之为碳源。能被微生物用作碳源的物质种类极其广泛,简单的无机含碳化合物(CO2、NaHCO3和CaCO3等)、比较复杂的有机物(烃类、醇类、羧酸、脂肪酸、糖及其衍生物、杂环化合物、氨基酸和核苷酸等)、复杂的有机大分子(蛋白质、脂类和核酸等),乃至复杂的天然含碳物质(牛肉膏、蛋白胨、花生饼粉、糖蜜、石油及其不同的馏分等)都可以被不同的微生物利用。甚至像二甲苯、酚等有毒物质均可以被少数微生物用作碳源。不同营养类型微生物利用不同的碳源。为数众多的异养微生物常利用某类有机化合物中的一种或几种物质作为它们的碳源。 | ||
| + | :[[糖的分类|糖类]]是微生物利用最广泛的碳源,尤其是葡萄糖。其次是醇类、有机酸和脂肪酸等。 | ||
| + | |||
| + | *氮源 | ||
| + | :氮是微生物细胞需要量仅次于碳的元素。能提供微生物生长代谢所需氮元素的营养物质称为氮源。能被微生物用作氮源的物质种类也很广泛:有分子态氮、氨、铵盐和硝酸盐等无机含氮化合物;尿素、氨基酸、[http://baike.baidu.com/item/%E5%98%8C%E5%91%A4 嘌呤]和[http://baike.baidu.com/link?url=7EK_uWzpa8wdJ4Jb7BaG69mECt5hE-cTg3xirIMY72XyBHqND3ACdCTDagqKlEAgC0y8Xi06gX54Z4b0sVsbiQeysj2Of29c2uNiK-EyQ5i 嘧啶]等有机含氮化合物。 | ||
| + | |||
| + | *无机盐 | ||
| + | :微生物还需要很多其他元素,如P、S、K、Mg、Ca、Na和Fe等大量元素,以及Co、Zn、Mo、Cu、Mn、Ni和W等微量元素。这些元素大多是以无机盐的形式提供的。其中,Mg、K、Na、Fe、Co、Zn、Mo、Cu、Mn和Ni等金属元素来源于无机盐的阳离子,而P、S等非金属元素绝大多数来自于无机盐的酸根。 | ||
| + | :无机盐的需要量虽然远小于C、N,但其重要性并不亚于它们。它们的生理功能可归纳为:提供微生物细胞化学组成中的重要元素,如P和S分别为核酸与含硫氨基酸的重要组成元素;参与并稳定微生物细胞的结构,如P参与的磷脂双分子层构成了细胞膜的基本结构,Ca参与细菌芽孢结构的皮层组成,Mg有稳定核糖体和细胞膜的作用;与酶的组成和活力有关。如Fe是细胞色素氧化酶的必要组分,Mg、Cu和Zn等是许多酶的激活剂;调节和维持微生物生长过程中诸如渗透压、氢离子浓度和氧化还原电位等生长条件,如Na和K有调节细胞渗透压的作用,由磷酸盐组成的缓冲剂能保持微生物生长过程中ph值的稳定,含S的Na2S和含疏基的疏基乙酸、半胱氨酸、谷胱甘肽和二硫苏糖醇等可降低氧化还原电位;用作某些化能自养型细菌的能源物质;用作呼吸链末端的氢受体。 | ||
| + | |||
| + | *生长因子 | ||
| + | :许多微生物除了上述的物质以及能源之外,还必须在培养基中补充微量的有机营养物质才能生长或者生长良好,这些微生物生长所不可缺少的微量有机物质就是生长因子。生长因子有维生素、氨基酸、嘌呤碱和嘧啶碱、卟啉及其衍生物、固醇、胺类、C2~C6直链或分支脂肪酸等。一些特殊的辅酶也能用作生长因子。能提供生长因子的天然物质有酵母膏、蛋白胨、麦芽汁、玉米浆等等。 | ||
| + | :生长因子的主要功能是提供微生物细胞重要化学物质(蛋白质、核酸和脂质)、辅助因子(辅酶和辅基)的组分和参与代谢。 | ||
| + | |||
| + | *水 | ||
| + | :水在细胞中的存在形式分为结合水与自由水:结合水是细胞结构的重要组成成分,用以保持生物活性(种子细胞中的结合水如果失去,那么将不再可发芽);自由水如下所述(失去自由水只要再次吸水后仍可在发芽)。 | ||
| + | :水是微生物营养中不可缺少的一种物质。这并不是由于水本身是营养物质,而是应为水是微生物细胞的主要化学组成:水是良好的溶剂;水具有运输物质的作用;水本身参与许多化学反应;水是良好的热导体,保证细胞内的温度不会因为代谢过程中能量的释放而骤然上升;水还有利于生物大分子结构的稳定; | ||
酵母的繁殖和生长可划分为六个不同阶段。 | 酵母的繁殖和生长可划分为六个不同阶段。 | ||
| 第248行: | 第308行: | ||
*矿物质的新陈代谢和生长因子 | *矿物质的新陈代谢和生长因子 | ||
:酵母的新陈代谢还取决于足够的矿物质和生长因子,这些物质的作用不可低估。部分离子对酶促反应影响很大,如钾离子与ATP仪器促进所有的酶促反应,对于能量代谢和细胞壁的物质输送很重要;钠离子使酶活化,在细胞膜的物质输送中起重要作用;钙离子可以被锰离子、镁离子所取代,延缓酵母退化,促进凝固物的形成;镁离子对有磷参与的反应十分重要,特别是发酵中不可取代;钙离子很少量就会抑制某些酶;铁离子对酶的呼吸代谢很重要,可促进酵母出芽繁殖;锰离子在代谢中可取代铁离子,可促进细胞繁殖和细胞形成;锌离子有利于蛋白质的合成,其需求量为0.2mg/L麦汁,缺锌可使发酵出现问题;硝酸根离子可被细菌还原为亚硝酸根,对细胞有毒性,极不利于发酵。 | :酵母的新陈代谢还取决于足够的矿物质和生长因子,这些物质的作用不可低估。部分离子对酶促反应影响很大,如钾离子与ATP仪器促进所有的酶促反应,对于能量代谢和细胞壁的物质输送很重要;钠离子使酶活化,在细胞膜的物质输送中起重要作用;钙离子可以被锰离子、镁离子所取代,延缓酵母退化,促进凝固物的形成;镁离子对有磷参与的反应十分重要,特别是发酵中不可取代;钙离子很少量就会抑制某些酶;铁离子对酶的呼吸代谢很重要,可促进酵母出芽繁殖;锰离子在代谢中可取代铁离子,可促进细胞繁殖和细胞形成;锌离子有利于蛋白质的合成,其需求量为0.2mg/L麦汁,缺锌可使发酵出现问题;硝酸根离子可被细菌还原为亚硝酸根,对细胞有毒性,极不利于发酵。 | ||
| + | |||
| + | ===影响酵母生长的因素=== | ||
| + | 微生物的生长受到它们所处环境因素的影响极大。微生物可能在某些有害条件下不能声张,但却可以忍受而不至于死亡,因而必须区分环境条件对微生物存货的影响与对微生物生长、分化和繁殖的影响之间的差别。比如某些条件下只会降低微生物的活性,但是达到临界点后完全永久性失活。 | ||
| + | |||
| + | *温度 | ||
| + | :温度是影响微生物生长的一个重要因子。温度太低,可使原生质膜处于凝固状态,不能正常地进行营养物质的运输或质子梯度,因而生长不能进行。当温度升高到适宜温度时,由于细胞内酶的催化作用使得化学反应以较快的速度进行,从而生长速率加快。 | ||
| + | :然而,当超过临街温度时,蛋白质、核酸和细胞其他成分就会发生不可逆的变性作用。因此,当温度在给定范围内升温时,代谢和生长就会加速,当超过临界点时完全失活。每种微生物都有3中基本温度,即最低生长温度、最适生长温度和最高生长温度。根据微生物的生长温度范围,可将其分为嗜冷微生物、嗜温微生物、嗜热微生物和嗜高热微生物。 | ||
| + | :由于酵母菌种的不同,每种酵母都有自己独立的最适温度,一般而言上面酵母(ALE)的最适温度要高于下面酵母(LAGER),具体的酵母温度应当参考所购酵母厂商给出的指标进行控制。 | ||
| + | |||
| + | *ph值 | ||
| + | :ph影响微生物的生长。因为它影响环境中营养物质的可给态和有毒物质的毒性;影响菌体细胞膜的带电荷性质、膜的稳定性以及膜对物质的吸收能力;使菌体表面蛋白变性或水解;酶的活性;每种生物都有一个可生长的ph值范围,以及最适生长ph值。大多数自然环境ph为5~9,适合于多数微生物的生长。 | ||
| + | :一般而言,啤酒酵母厂商没有给出最适ph。啤酒的ph在糖化阶段一般控制在5.2左右,即使到发酵后期ph下降后,各类型的酵母均能很好的生存并工作。所以只要控制好酵母投放时麦汁的ph值,不需要过于担心ph对酵母带来的影响。 | ||
| + | |||
| + | *氧 | ||
| + | :微生物对氧的需要和耐受能力在不同的类群中变化很大,依据它们和氧的关系可分为几种类群。 | ||
| + | ::*好氧微生物 | ||
| + | :::包括所有需要氧才能生长的微生物。有两类:一类是专性好氧微生物,它们的生长必须要有氧,快速分裂的细胞比缓慢分裂的细胞需要的氧更多,通常生长在培养基表面附近;另一类是微好氧微生物,它们在有少量自由氧存在条件下生长最好,因而生长在培养基表面之下的某一区域,该区域氧浓度正好符合它们生长的需要。 | ||
| + | |||
| + | ::*厌氧微生物 | ||
| + | :::那些缺乏呼吸系统而不能利用氧气作为末端电子受体的微生物称之为厌氧微生物。可分为两类:耐氧厌氧微生物和严格厌氧微生物。前者是指那些尽管不需要氧,但可耐受氧,并在氧存在条件下仍能生长的类群;而后者则是指那些对氧敏感,在有氧时既被杀死的类群,所以专性厌氧微生物只能生长在氧气几乎不能达到的培养基底部附近,严格厌氧微生物并不是被气态的氧所杀死,而是由于不能解除某些氧代谢产物的毒性而死亡。 | ||
| + | |||
| + | ::*兼性好氧微生物 | ||
| + | :::在有氧存在下通常进行有氧呼吸,产生CO2和水以及大量的能量,但在氧缺乏时可以转变为无氧呼吸,产生酒精、乳酸等代谢物和CO2以及少量的能量。这类微生物在有氧呼吸条件下的生长必无氧条件下的生长更旺盛,因而可以看到菌体在整个培养基中都有分布。 | ||
| + | :::酵母菌属于兼性好氧微生物,在有氧条件下进行快速的生长繁殖并产生出大量的热和少量的CO2,在无氧条件下产生酒精和CO2。 | ||
=啤酒酵母和啤酒质量的关系= | =啤酒酵母和啤酒质量的关系= | ||
在啤酒生产中酵母菌体不是最终产品,但对最终产品的质量非常重要,发酵过程中一系列的复杂生化反应均系酵母营养代谢作用而致,故酵母对啤酒生产和发酵质量,乃至啤酒的理化性能和其风味典型性,均有重要影响。 | 在啤酒生产中酵母菌体不是最终产品,但对最终产品的质量非常重要,发酵过程中一系列的复杂生化反应均系酵母营养代谢作用而致,故酵母对啤酒生产和发酵质量,乃至啤酒的理化性能和其风味典型性,均有重要影响。 | ||
| − | * | + | ===酵母与发酵速度=== |
| − | : | + | 酵母的发酵速度是啤酒生产的重要指标之一。在一定的工艺技术条件下,酵母对麦汁顺利地完成发酵,为啤酒的质量提供了保证。同时,在保证发酵质量的前提下,充分发挥酵母细胞内在的潜力,可加速生产周期的循环,提高生产能力。 |
| − | : | + | 影响酵母发酵速度的因素主要有以下几个方面 |
| + | *酵母浓度 | ||
| + | :在充分搅拌并使酵母细胞均匀分布于发酵液中的情况下,发酵速度与酵母的浓度成正比例关系,如: | ||
| + | ::接种量0.6L/hl,发酵天数为9天; | ||
| + | ::接种量1.0L/hl,发酵天数为7天; | ||
| + | ::接种量2.0L/hl,发酵天数为4~5天; | ||
| + | :当然,在实际生产中无需采用过高的酵母接种量,因为过高接种量容易使酵母衰退,发酵现象也不易控制。 | ||
| + | |||
| + | *酵母发酵力 | ||
| + | :发酵力用来衡量酵母酒精发酵的能力,因为环境的影响而有很大变化,如酵母的贮存条件、细胞表面的附着物等对发酵力均有影响。而酵母的菌龄、菌株性质的不同,其发酵力也不同。酵母代谢产物的原生质毒素,也会抑制其发酵力。当发酵生成酒精超过8.5%时,发酵也会被抑制,今儿降低酵母的发酵力。 | ||
| + | |||
| + | *麦汁组成 | ||
| + | :麦汁中的铜被酵母吸收累积至一定程度,将使酵母衰退而减缓发酵速度;麦汁中可被同化氮源的含量也影响发酵速度,一般情况是发酵力随含氮量的增加而提高;酵母在麦汁中发酵糖分的最适合PH值是4~6,超出范围将对发酵速度产生影响。 | ||
| + | |||
| + | *发酵温度 | ||
| + | :温度将直接影响酵母的繁殖和发酵力,尤其是下面酵母,较高的温度能加快发酵速度。 | ||
| + | |||
| + | ===酵母与发酵度=== | ||
| + | 啤酒酵母的发酵度反应酵母对各种糖类的发酵情况。正常的啤酒酵母能发酵葡萄糖、果糖、蔗糖、麦芽糖和麦芽三糖。酿制不同类型的啤酒,需要不同的发酵度。有的酵母具有较高的发酵度,有的酵母不可发酵麦芽三糖而使发酵度降低。 | ||
| + | |||
| + | 在一些非正常情况下,如果酵母的发酵度降低,一方面说明酵母有变异或污染的可能,另一方面应检查麦汁成分及发酵条件是否恰当。'''发酵度决定了啤酒类型和口味,一般控制啤酒发酵度:外观发酵度为65%~80%,真正发酵度为55%~70%。一般来说,发酵度低的啤酒并不醇厚,只是黏口、腻厚和甜感,其保质期也短;高发酵度的啤酒多数醇厚,具备了啤酒的“酒体”'''。 | ||
| + | |||
| + | ===酵母与发酵异常现象=== | ||
| + | 在啤酒的发酵过程中,常常遇到一些发酵异常现象,包括主发酵和后发酵期间的异常现象 | ||
| + | |||
| + | ====主发酵期间的发酵异常现象==== | ||
| + | =====裂纹现象===== | ||
| + | 在主发酵期间的起泡期和高泡期,发酵液表面布满泡沫时,发生液面泡沫开裂,泡沫慢慢变薄,而且不均匀,发酵不旺盛。发生这种现象的主要原因:一方面是洗涤酵母后,贮存室水温和室温变高,促进酵母代谢作用加强而缺氧,酵母衰老,造成发酵减退;另一方面是由于麦汁中α-氨基氮含量不足,溶解氧含量少,接种温度低,以及麦汁浑浊使酵母细胞表面吸附过多的蛋白质和酒花树脂,酵母的酶不能与糖类作用,使发酵变为迟缓,从而出现了裂纹现象。 | ||
| + | |||
| + | 防止和解决办法:在糖化时用乳酸或磷酸调整醪液的ph值;延长和促进蛋白质休止时间;提高麦芽汁α-氨基氮含量;提高麦芽汁接种温度和麦芽汁中的氧气含量;提高酵母使用量。 | ||
| + | |||
| + | =====泡沸现象===== | ||
| + | 泡沸发酵也称为沸腾发酵。常在主发酵后期或落泡期或下酒捞出泡沫时出现,一种现象是发酵液表面的泡盖由一角或一边推向另一边,部分页面又出现白色泡沫;另一种现象是大量的二氧化碳气泡上涌,发酵液像喷泉一样剧烈翻动,把以沉淀的酵母块带到液面。在这种现象未发生前,朱发酵现象如降糖都是正常的。 | ||
| + | |||
| + | 泡沸发酵是时有发生的,但发生的原因还不是十分清楚,说法很多,其中比较合理的解释是:一是酵母不纯,有产生气体的微生物,随酵母沉积到底部,产生大量的二氧化碳,积聚到沉淀的酵母中,最后把酵母层冲开,急剧上升,形成沸腾;二是麦汁组成成分不良,麦汁浑浊不清,固体随酵母沉淀到底部,酵母继续利用固体的营养物质发酵产生二氧化碳,积聚在其中,量大时把酵母冲开,二氧化碳气体随之上升而沸腾;再一种是主发酵温度过高,后期采取急剧降温,当把泡盖去掉后,表面压力降低,使下边二氧化碳急剧上升造成沸腾;还有一种是啤酒酵母变异,凝聚性不良,而且麦汁可发酵性糖比例偏高,产生旺盛发酵,二氧化碳产生量多,在泡盖捞出后,二氧化碳急剧上升所造成。 | ||
| + | |||
| + | 对于沸腾发酵,先在技术管理上还没有有效的办法解决,一般采取改进糖化操作,改善麦汁组成成分,加强麦汁过滤,使麦汁清亮透明,另外,从加强酵母管理着手,遇到泡腾发酵时,重新培养酵母,并加强卫生管理和灭菌工作。 | ||
| + | |||
| + | =====气泡发酵现象===== | ||
| + | 气泡发酵也称为异泡发酵,即在主发酵期的低泡期,或发酵终了前一天,发酵液表面已形成的棕色泡盖上出现多数的大气泡,或称为大明泡,继而破坏了泡盖,使已凝结出来的酒花树脂和蛋白质凝结物下沉,使表面的泡盖变成白色的现象。产生气泡发酵的主要原因是酵母无染杂菌、野生酵母或其他原因,其次是糖化不完全,糖化用水中亚硝酸盐过量。出现这种情况后发酵液不易澄清,啤酒的口味不纯。 | ||
| + | |||
| + | 防止的办法:加强酵母室和发酵室的卫生管理工作,及时做好清洁、灭菌工作和现场使用的酵母的洗涤保管工作,及时培养新鲜强装的酵母,按期更换,加强糖化管理,对糖化用水定期检测,受亚硝酸盐和硝酸盐污染的水要进行处理,根据原料情况调整糖化操作,使生成的麦汁符合要求。 | ||
| + | |||
| + | =====虚泡现象===== | ||
| + | 在主发酵落泡期,形成疏松的泡沫,开始是白色,逐渐变成棕黄色,最后泡盖松散无力,凝结在下面的酒花树脂沉入发酵液内。这主要是由于原料麦芽溶解不好,糖化时蛋白质分解的温度和时间不恰当所造成。 | ||
| + | |||
| + | 防止的办法:对原料事先做好监测工作,根据原料情况制定糖化工艺操作方法,加强半成品的分析。 | ||
| − | + | =====发酵中止现象===== | |
| − | + | 主发酵达到高泡期,泡沫升起不久,很快又回缩,糖度下降缓慢,甚至出现发酵中止现象,同时发酵液澄清。产生发酵中止现象的原因是多方面的,如糖化所得的麦汁α-氨基氮或微量物质嘌呤、嘧啶含量不足,可发酵性糖含量过低,糊精等非糖含量过高;糖化时醪液的PH值过高,使植酸钙和镁盐不能充分分解为肌醇和磷酸盐,使麦汁中缺乏生长素;麦汁中的酸度过高或过低,极易造成酵母沉淀;在主发酵时,温度掌握不当,突然降低温度,使酵母过度受刺激而沉淀;酵母发生变异,不能发酵麦芽三糖等。发酵中止可造成发酵液发酵度低、残糖高、有甜味、口味淡薄、不爽快、泡持性不好。 | |
| − | + | 解决的办法:好次原料搭配使用,防止糖化麦汁质量不一,同时在糖化时,对糖化用水加强处理,调整水的PH值,有利于提高可发酵性糖和可同化氮化物的含量,以及生长素的含量,在糖化时促进蛋白质的分解和麦汁煮沸时的凝固;严格管理发酵工艺,防止温度忽高忽低,避免酵母受刺激。发生这种现象后可采取倒桶、添加酵母、通风搅拌、供应充足的氧气,使之重新发酵;也可将中止发酵的发酵液倒入两个发酵罐中,分别加满已繁殖后的发酵液,使之重新发酵。 | |
| − | + | ||
| + | =====再发酵现象===== | ||
| + | 在主发酵末期,泡盖已经形成,忽然又开始旺盛发酵,形成白色泡沫,将已形成的棕色泡盖翻入发酵液中。发生这种现象的主要原因是,麦汁组成成分不当,可发酵性糖少,但糊精的中间产物通过酵母中酶的作用,又被酵母所利用,另一种原因是酵母变异了。 | ||
| + | |||
| + | 解决的办法是对所使用的酵母要做性质检查,确定酵母是否变性,目前有些厂所使用的酵母经过长期的高温发酵,降糖速度很快,一天可以降糖4~5°P,这也是酵母变性问题,另一种解决办法是调整糖化工艺。 | ||
| + | |||
| + | ====后发酵期间的发酵异常现象==== | ||
| + | =====发酵不旺盛现象===== | ||
| + | 发酵不旺盛,开口发酵时造成泡沫不溢出的原因主要有:下面发酵液中酵母细胞少;发酵罐中所留空隙太大;酵母衰老,发酵作用已极为微弱。解决措施:可采用添加高泡酒的方法。 | ||
| + | |||
| + | =====贮酒罐不升压现象===== | ||
| + | 贮酒罐封罐后,在3~10天内罐压应升至0.05~0.08MPa,若封罐后不升压应检查原因,如贮酒罐是否漏气;酒液中酵母细胞数是否过少;一般可采取倒罐、加高泡酒的方法进行解决。 | ||
| + | |||
| + | =====发酵沉淀不清现象===== | ||
| + | 贮酒较长时间后,酒液浑浊不清,其原因:糖化不完全,蛋白质分解欠佳;麦汁中α-氨基氮过少;PH值不当;酵母被杂菌污染。 | ||
| + | |||
| + | 解决措施:应重新调整糖化工艺,调整麦汁组成,或倒罐至其他新发酵的酒液罐内;若酵母感染杂菌,则应该处理掉。 | ||
=酵母的检查与鉴定= | =酵母的检查与鉴定= | ||
| + | 啤酒酵母的质量直接关系到啤酒发酵和啤酒的质量。如果啤酒酵母被杂菌污染或发生变异,就会产生不正常的发酵现象和影响啤酒的口味。酵母的自然变异是比较低的,但是在长期的酵母培养和发酵过程中变异的可能还是存在的。 | ||
| + | |||
| + | ==啤酒生产中酵母的检查== | ||
啤酒生产过程中,经常对酵母进行镜检和做某些生理特性试验,镜检一般只起辅助作用,对酵母某些生理特性的检查更具有重要性。 | 啤酒生产过程中,经常对酵母进行镜检和做某些生理特性试验,镜检一般只起辅助作用,对酵母某些生理特性的检查更具有重要性。 | ||
| − | + | ===外观和形态检查=== | |
| − | + | ====菌落形态==== | |
| − | : | + | *液体培养基中观察菌落形态 |
| + | :在液体培养基中观察发酵液浑浊的快慢,澄清的程度及酵母沉淀的情况。酿造车间现场使用的酵母泥必须新鲜,呈黄白色,有果实的爽快香味,其上部洗涤水透明且无色。沉于底部的酵母泥紧密,取起后应松散而不黏连。如色泽深暗或发粘则说明质量较差。 | ||
| − | : | + | *啤酒酵母个体形态观察 |
| + | :用显微镜观察酵母细胞的形状、大小、夹杂物以及是否有细菌等。啤酒酵母呈球形、椭圆形或卵圆形,细胞的平均直径为4~5μm,大小为(3~7)μm×(5~10)μm。液体培养的酵母细胞大于固体培养的细胞。成熟细胞大,年幼时细胞小。 | ||
| − | : | + | :优良健壮的酵母细胞具有均匀的形态和大小,平滑而薄的细胞膜,细胞质透明均一,年幼少壮的酵母细胞内部充满细胞质;老熟的细胞出现空泡,内贮细胞液,呈灰色,折光性强;衰老的酵母死亡率高,可通过美蓝染色,检查酵母死亡率。一般生产上使用的酵母死亡率应在3%以下,新培养的酵母死亡率在1%以下。 |
| − | |||
| − | : | + | :下面发酵啤酒酵母一般以单端出芽繁殖,很少形成短链。芽在脱离母细胞前总是比母细胞小,芽和母细胞的纵轴有30°夹角,这和许多野生酵母的芽和母细胞在一个纵轴上形成鱼鳔形有明显区别。在镜检中,如果发现显著变异,可怀疑是酵母退化或可能有杂菌污染,需要另做较细致的细菌或野生酵母鉴定。夹杂物为蛋白质和酒花树脂等,如与细菌等分辨不清时,可在酵母中加入10%碱溶液或50%醋酸,则蛋白质小颗粒溶解,标本中只看到酵母和细菌。 |
| − | + | ====巨大菌落的观察==== | |
| + | 某些酵母的品种鉴别上,在一般的形态上不易区别。但是,它们所生成的巨大菌落则不一样,菌落越大形态越容易区别。一般,巨大菌落表面平滑多为分散型酵母,而表面褶皱都为芽簇型酵母。巨大菌落和发酵性能之间没有什么联系。有的菌种巨大菌落为白色,表面平滑呈扁平或半透明镜状隆起,有时中部略呈凹形。 | ||
| − | + | ====杀菌及其污染程度的观察和检查==== | |
| − | + | 确定50个[[显微镜视野]]中存在的杂菌数并按等级确定污染程度: | |
| − | + | {| class="wikitable" | |
| − | + | |- | |
| + | | 杂菌数 || 污染等级 | ||
| + | |- | ||
| + | | 1个 || 微量 | ||
| + | |- | ||
| + | | 3个 || 很少 | ||
| + | |- | ||
| + | | 6个 || 少 | ||
| + | |- | ||
| + | | 8个 || 轻度污染 | ||
| + | |- | ||
| + | | >8个 || 强度污染 | ||
| + | |} | ||
| − | + | 对单一微生物允许污染标准如下: | |
| + | {| class="wikitable" | ||
| + | |- | ||
| + | | 野生酵母 || 很少感染(3个) | ||
| + | |- | ||
| + | | 细菌(杆菌、乳酸菌、四链球菌) || 少感染(6个) | ||
| + | |- | ||
| + | | 无损害的微生物(球菌和小酵母) || 中等感染 | ||
| + | |} | ||
| − | + | 按照以上标准,在每次酿造前应该精确检查,并确定污染程度和新细胞的比率。应该保持一个视野中约有100~200个酵母细胞,因此悬浊液是较浓的。 | |
| − | + | ====死细胞的检查==== | |
| − | + | 良好的现场使用酵母其死细胞数一般在0.5%~3%之间,不应超过5%。检查时取磷酸缓冲液(0.2mol/L磷酸氢二钠0.25ml和0.2mol/L磷酸二氢钾99.75ml混合)和0.04%美蓝溶液等量混合,既得0.02%美蓝染色液,其ph为4.6,此溶液需保存于暗处。同时取适量浓度的酵母悬浊液1ml(泥状酵母1ml用水稀释成200倍),混合1ml的染色液,5min内在显微镜下检查,数出被染色(蓝色)的酵母死细胞数,计算其百分率,即为酵母细胞死亡率。 | |
| − | + | ====肝糖染色检查==== | |
| − | + | 发酵力强的酵母细胞始终含有肝糖。肝糖在强盛的发酵阶段形成,其数量决定于麦芽汁的组成。如酵母长期在水下保存(3~5天)则肝糖完全消失,因为肝糖作为贮存物质一部分被酵母所消耗,同时被酶催化分解后的一部分也转移到水中。不旺盛的发酵或者发酵开始迟缓都是肝糖成分不足的标志。一般肝糖都是在纯培养时进行测定,该测定项目也可以在生产中发现发酵不良时进行。但是目前还不能精确地指出一个发酵力强的酵母应该含有染成怎样程度的褐色细胞。肝糖含量首先决定于酵母的生理形态,其次取决于生长的强度和贮存时间。发酵后取出的健康教母应有70%为染成深褐色的细胞。 | |
| − | |||
| − | + | ====异染颗粒的检查==== | |
| − | + | 新鲜强装的酵母异染颗粒粒大且色深,根据其含量可以判断酵母在生理学上的能力。很多学者认为,异染颗粒的含量与酵母的发酵能力和繁殖之间存在着一定的关系。异染颗粒是根据基质中磷酸化合物的存在而存在,在判断老的和退化的酵母时,对其进行检查是可行的。 | |
| − | + | ====芽蔟的检查==== | |
| + | 将现场使用的酵母稀释并制成悬滴镜片,在显微镜下观察酵母的连结是由于凝聚力还是由芽蔟形成所致。如果识别有困难,可在酵母中加稀薄的氢氧化钾溶液或稀醋酸溶液,然后再进行显微镜检查。此时,凝集酵母细胞即个个分离而芽蔟细胞仍在。下面发酵现场使用的酵母如果有芽蔟存在,可以人为是由于异种酵母的混入所致。 | ||
| − | + | ====孢子形成速度的检查==== | |
| − | : | + | 将酵母移植到酒石酸蔗糖溶液中(酒石酸4g,蔗糖10g,溶于100ml水中),20℃培养48h,如此进行2~3次,再在麦汁液体培养基中繁殖24h后移植到石膏块上(石膏块应先刮平,上有凹部,放入双重皿中杀菌,以水浸没石膏块1/2处),在25℃恒温箱中放72h,用显微镜检查孢子形成情况。如是野生酵母,此时已经形成孢子。 |
| − | + | ||
| − | + | ==酵母生理特性试验== | |
| − | | | + | ===发酵力=== |
| + | 酵母发酵力用以判别其酒精发酵的能力,一般来说应该选择发酵能力强的酵母。如果酵母发酵力衰退则意味着酵母发生退化、变异。发酵力测定的方法有以下两种。 | ||
| + | |||
| + | ====发酵法==== | ||
| + | 取灭菌的250ml三角瓶,加上棉塞,瓶内加入150ml麦汁,经过常压灭菌,冷却后加入酵母泥1g摇匀,放在25℃保温箱中进行发酵,每隔8h震动1次,经过3~4天发酵终了,过滤掉酵母,取发酵液100g,并把酒精蒸馏出去,放在定量瓶中添加蒸馏水使重量恢复到100g,混合摇匀,测量20℃时的密度,测出残留在发酵液中的浸出物浓度,利用下面的公式计算真正发酵度: | ||
| + | |||
| + | :真正发酵度(%)=(发酵前麦汁浓度 - 发酵后排除酒精的麦汁浓度) / 发酵前麦汁浓度 | ||
| + | |||
| + | 发酵后,不用蒸馏法去除酒精,直接测量密度,算出发酵液残留浸出物的浓度,利用下面的公式计算其外观发酵度: | ||
| + | |||
| + | :外观发酵度(%)=(发酵前麦汁浓度 - 发酵后麦汁浓度) / 发酵前麦汁浓度 | ||
| + | |||
| + | 外观发酵度一般比真正发酵度高约10%,换算方法如下:真正发酵度 = 外观发酵度×0.819。啤酒的发酵度一般都分为高、中、低三个类别,见下表: | ||
| + | {| class="wikitable" | ||
|- | |- | ||
| − | | | + | | rowspan="2" | || colspan="2" | 淡色啤酒 || colspan="2" | 浓色啤酒 |
| − | | | ||
|- | |- | ||
| − | | | + | | 外观发酵度 % || 真正发酵度 % || 外观发酵度 % || 真正发酵度 % |
| − | | | ||
| − | | | ||
|- | |- | ||
| − | | | + | | 低发酵度 || 60 - 70 || 48 - 56 || 50 - 58 || 41 - 47 |
| − | | | ||
| − | | | ||
|- | |- | ||
| − | | | + | | 中发酵度 || 73 - 78 || 59 - 63 || 60 - 66 || 48 - 53 |
| − | | | ||
| − | | | ||
|- | |- | ||
| + | | 高发酵度 || > 80 || > 65 || > 70 || > 56 | ||
|} | |} | ||
| − | + | ====二氧化碳减量法==== | |
| − | + | 用已知重量的250ml发酵瓶装入12 ~ 18°P的麦汁150ml,在101.325Pa下灭菌15~20min,冷却后添加酵母泥1g,接上发酵栓(类似水封,里面加入杀菌液可以洗涤气体),擦干瓶外的水汽,称其重量。把发酵瓶放在20℃的保温箱中发酵,每天定时称重,发酵6~8天,最后每天减重不高于0.2g,即为发酵终止。 | |
| − | + | ||
| + | 优良的酵母发酵力强,二氧化碳气体溢出多,一般中等发酵的酵母在12°P麦汁中发酵失重> 3.6g,若逐日比较二氧化碳失重情况,由此可判别发酵速度。 | ||
| + | |||
| + | ===酵母热死亡温度=== | ||
| + | 微生物的热死亡温度是指液态培养的微生物,在某温度下即刻被杀死,此温度称为微生物的热死亡温度。啤酒酵母一般在45℃时即停止生命活动,热死亡温度为50~54℃。 | ||
| + | |||
| + | 为了避免试验酵母的缺点,习惯上先将试验酵母移到液体培养基中在25℃培养24h后试用,或从扩培中采取以供实验。酵母的热死亡温度除了与培养基种类有关外,与加热时间长短也有关。啤酒厂选择的温度为40~60℃,每个间隔温度为2℃,保温时间习惯以10min为度。 | ||
| + | |||
| + | 若酵母的热死亡温度改变,说明菌种发生变异,或受到野生酵母污染。野生酵母比培养酵母有更高的耐热性。 | ||
| + | |||
| + | ===酵母的凝聚性试验=== | ||
| + | 啤酒酵母的凝聚性是区别菌株的一项重要内容,在生产商具有特殊的重要性。各种酵母的凝聚性有较大的差别,当酵母发生变异或衰老时,凝聚性随之发生较大的变化。啤酒酵母的凝聚性不同,酵母的沉淀速度也有差异,发酵也不一样。凝聚性强的酵母,发酵液容易澄清,但发酵度偏低;凝聚性差的酵母,发酵液不易澄清,酵母回收困难,但是发酵度高。 | ||
| + | |||
| + | 啤酒酵母凝聚性测定的方法采用本斯(Burns)实验法:将1g酵母泥与10ml、pH4.5的醋酸缓冲液混合,20℃平衡20分钟,加至带刻度的锥形离心管内,连续20分钟,每隔1分钟记录沉淀酵母的容量。实验后,检查pH是否保持稳定。一般规定10分钟时的沉淀酵母量在1.0ml以上者为强凝集性,0.5ml以下者为弱凝集性。 | ||
| + | |||
| + | ===发酵速度的测定=== | ||
| + | 发酵速度与酵母品种有关,如酵母的麦芽糖酶活性是控制麦芽糖发酵的重要因素,与发酵速度关系很大;发酵速度与环境条件的关系也很密切,如麦汁成分、发酵温度、通风条件、发酵容器等。一般来说酵母在统一条件下发酵速度越快越好。 | ||
| + | |||
| + | 为了取得与现场发酵条件相似的发酵速度,测定方法是:在直径5cm、长120cm的玻璃筒内,装2L麦汁,接种后按现场发酵条件控制,每天测定其外观浓度,观察对比其发酵速度。 | ||
| + | |||
| + | ===感官鉴定=== | ||
| + | 不同的啤酒酵母,其发酵时的代谢产物不尽相同,因而发酵液的风味也不一样。只有优良的啤酒酵母才能产出优秀风味的啤酒,不仅风味好,还要有正常的芳香,而且要求风味始终保持一致,如果生产过程中产生怪味和异味,就必须检查所用酵母是否发生变异或污染。 | ||
| − | + | ===耐酒精浓度的试验=== | |
| − | + | 酵母在麦汁中发酵,到某一程度即停止。其原因,一是由于可发酵性糖的耗尽,二是受酒精含量的抑制。这在实际应用上具有很大意义。虽然在通常的啤酒发酵中,酒精含量较低,对酵母的影响不大,但不同的酵母对酒糟浓度的耐受力各有不同,在发酵时,一般采用能耐受较高酒精度的酵母,以有利于发酵。 | |
| − | |||
| − | |||
| − | + | ===染色实验=== | |
| − | + | 通过[[美蓝染色]]试验计算其死亡率。新酵母(包括或扩培后的酵母)的死亡率应当< 1%,现场使用中的酵母死亡率应当< 3%。 | |
| − | + | ===降糖速度=== | |
| − | + | 啤酒酵母的发酵能力可用降糖速度来表示。正常培养的酵母,第10天外观浓度应该不高于3.5°P,如果在4.0°P以上则为降糖慢的酵母。 | |
2017年1月6日 (五) 19:03的最新版本
麦汁经过啤酒酵母的发酵作用后,便酿制成为啤酒。由于酵母不仅进行酒精发酵,而且其新陈代谢的产物还影响啤酒的口味和特点,所以酵母也被称为啤酒的灵魂,了解酵母的结构和组成、新陈代谢、繁殖和生长及其分类非常重要。不同的酵母菌种有一系列不同的特性。
啤酒酵母的特性
啤酒酵母在分类学上的位置
按照微生物分类系统,啤酒酵母属于真菌门,子囊菌纲,原子囊菌亚纲,内孢霉目,内孢霉科,酵母亚科,酵母属,啤酒酵母。
工业生产上应用的酵母菌都属于酵母属,在自然界分布很广,有很多菌种,其中以啤酒酵母最为重要。啤酒酵母种类较多,不同品种的菌株,在形态以及生理上都有明显的区别。
啤酒酵母的命名
啤酒酵母是根据国际命名法则命名的。啤酒酵母的学名为Saccharomyces cerevisiae,拉丁语cerevisiae意为麦酒。酵母是属名,习惯翻译,以种名放在前面,属名放在后面。
啤酒酵母又分为上面发酵酵母(ALE)和下面发酵酵母(LAGER)。汉生(Hansen)首先从苏格兰爱丁堡啤酒厂分离出上面发酵的纯粹培养酵母,此类啤酒酵母即命名为Saccharomyces cerevisiae Hansen。而后,汉生又在丹麦卡尔斯伯啤酒厂分离出下面发酵的纯粹培养酵母,命名此类啤酒酵母为Saccharomyces carlsbergensis Hansen。许多菌种,在种内包括有变种。表示变种的学名,是在该细菌学名后面加变种名称,并在变种名称前加var.(variety),意即变种。如啤酒酵母浑浊变种的学名为Saccharomyces cerevisiae var.turbidans。
酵母菌的分类
- 分类原则
- 酵母的分类较复杂,除了根据其形态特征外,还必须结合其生理生化特征。
- 形态特征
- 包括观察酵母在麦汁琼脂上的菌落形态、颜色、质地,在麦汁培养基中产醭、菌环、沉淀等外观,以及酵母在麦汁中及载片培养时营养细胞的形态、大小、繁殖方法、子囊孢子的形态及大小等。、
- 生理生化特征
- 包括发酵各种糖的能力,利用各种碳水化合物生长的能力,同化酒精的能力,能否利用硝酸盐,分解杨梅素,产生胡萝卜素,产生酯类,发酵牛奶,形成酸等。
- 酵母的分类系统过去很不一致,子层荷兰的酵母工作者娄德(Loder)于1952年发飙了酵母分类的系统著作之后,酵母的分类工作才较为一致。
- 培养酵母和野生酵母
- 培养酵母
- 啤酒厂中所使用的啤酒酵母属于有孢子真酵母菌,是由野生酵母经过系统、长期的驯化,并经过反复使用和考验,具有正常生理状态和特征,适合于啤酒工厂生产要求的培养酵母。
- 野生酵母
- 啤酒厂将凡是与培养酵母的形态和特征不一样,不为生产所控制利用的酵母称为野生酵母,如巴氏酵母,在自然界分布很广,存在于啤酒厂,妨碍啤酒的正常发酵,对啤酒生产有很大危害。
- 培养酵母和野生酵母的区别
- 单纯从外观形态上区别培养酵母和野生酵母是困难的,主要应从抗热性能、发酵糖类的性能、形成孢子的情况、在选择培养基上的生长情况等生理特征以及利用免疫荧光技术加以区别。
区别项目 培养酵母 野生酵母 细胞形态 圆形或卵圆形 有圆形、椭圆形、柠檬形等多种形态 抗热性能 在水中53℃,10分钟死亡 比培养酵母能耐较高的温度 孢子形成 较难形成 交易形成,有的野生酵母不形成孢子,但可以从细胞形状区别 糖类发酵 对葡萄糖、半乳糖、麦芽糖、果糖等均能发酵,能全部或部分发酵棉籽糖 绝大多数野生酵母不能全部发酵左旋的糖类。 含放线菌酮的培养基(0.2mg/L) 不能生长 非酵母属的野生酵母可耐受 以赖氨酸为唯一碳源的培养基 不能生长 非酵母属的野生酵母可生长 含结晶紫的培养基(20mg/L) 不能生长 酵母属的野生酵母可生长 棉衣荧光试验 可以区别 可以区别
- 上面酵母和下面酵母
- 上面酵母又称顶面酵母(ALE),啤酒厂使用的上面酵母是纯粹培养酵母;下面酵母又称底面酵母或储存酵母(LAGER),啤酒厂使用的下面酵母也是纯粹培养酵母。下面酵母是在不断变化的外界因素影响下,由上面酵母演变而来的。
- 形态和生理特征区别
区别项目 上面酵母(ALE) 下面酵母(LAGER) 细胞形态 多呈圆形,多数细胞集结在一起 多呈卵圆形,细胞较分散 发酵时的生理现象 发酵终了,大量细胞悬浮在液面 发酵终了,大部分细胞凝集在容器底部 芽细胞分支 生长培养时,生出有规则的芽细胞分支,易形成芽蔟 芽细胞分支不规则,且易分离,不易形成芽蔟 对棉籽糖发酵 能将棉籽糖分解为蜜二糖和果糖,只发酵三分之一的果糖部分 能全部发酵棉籽糖 对蜜二糖发酵 缺乏蜜二糖酶,不能发酵蜜二糖 含蜜二糖酶,能发酵蜜二糖 37℃培养 能生长 不能生长 孢子的形成 培养时相对较易形成孢子 很难形成孢子,只有用特殊培养方法才有可能 产生H2S或甲基硫醇 较低 较高 呼吸活性 高 低 对甘油醛发酵 不能 能 利用酒精生长 能 不能
- 其他区别
- 凝聚酵母和粉状酵母
- 凝聚酵母
- 啤酒酵母的凝聚性是酵母的生理特征之一,凝聚的强弱受基因控制极不一致。凝聚酵性强的酵母,从酒液中分离早,酒液中细胞密度低、沉淀快、发酵慢、发酵度低。凝聚性弱的酵母,与酒液分离晚,酒液中细胞密度高、沉淀慢、发酵快、发酵度高,回收酵母少、啤酒过滤困难。介于强弱之间的凝聚性则有较大的范围,酿造者应根据自己生产啤酒的类型进行选择。每一种菌种均有其一定的凝聚特点,在其他条件不变的情况下,凝聚性的改变往往是菌种变异的象征。
- 粉状酵母
- 粉状酵母又称絮状酵母。长时间悬浮在发酵液中不易沉淀,发酵结束时只有极少量松散的酵母沉淀为粉状酵母。上面酵母和下面酵母中均有粉状酵母。粉状酵母发较快、发酵度高,但回收困难,需用离心机回收酵母。
酵母种类 凝聚酵母 粉状酵母 发酵时的情况 酵母易于凝聚沉淀(下面酵母)或凝聚后浮在发酵液液面 不易凝聚 发酵终了 很快凝聚,沉淀密致,或于液面形成致密的厚层 长时间地悬浮在发酵液中,很难沉淀 发酵液澄清情况 较快 不易 发酵度 较低 较高 回收情况 容易回收 不易回收
酵母的形态与结构
酵母是一个俗称,一般泛指能发酵糖类的各种单细胞真菌。由于不同的真菌在进化和分类地位上的异源性,因此很难对酵母下一个确切的定义,通常认为,酵母具有以下五个特点:个体通常以单细胞存在;多数出芽繁殖;能发酵糖类产能;细胞壁含有甘露聚糖;常生活在含糖量较高、酸度较大的水、土环境中。
- 酵母的形态
- 自然界中的酵母形态以圆形为主,常见的有卵圆形、球形、柱形、腊肠形、瓶形、三角形。细胞大小一般在2.5~10μm × 4.5μm × 21μm。啤酒酵母呈圆形或卵圆形,细胞大小一般为3~7μm × 5~10μm。培养酵母的细胞平均直径为4~5μm,不能游动。啤酒酵母细胞形态往往受黄静影响而变化,但在环境好转后,仍可恢复原来的形态。啤酒酵母在麦汁固体培养基上,菌落呈乳白色、不透明,但有光泽,菌落表面光滑、润湿,边缘整齐。随着培养时间的延长,菌落光泽逐渐变暗,菌落一般较厚,易呗接种针挑起。啤酒酵母在液体培养基中,会在液体表面产生泡沫。常因菌种悬浮在培养基中而呈浑浊状。发酵后期,有的酵母悬浮在液面,形成一层厚膜,如上面发酵酵母;有的沉积于底部,如下面发酵酵母。
- 酵母细胞的结构
- 在显微镜下观察啤酒酵母细胞的结构,主要有细胞壁、细胞膜、细胞核、细胞质、液泡、线粒体等。

- 细胞壁
- 位于细胞的最外层,具有一定的弹性,决定着酵母细胞的形状和稳定性,约占细胞质量的30%,壁厚100~200nm。细胞壁由大分子的物质组成,主要成分为30%~40%的甘露聚糖(即酵母胶体)和30%~40%的葡聚糖。位于细胞外部的甘露聚糖与磷结合,而位于细胞里面的葡聚糖与硫以酯键连接,总复合物还包括蛋白质和酶,它们通过细胞膜分解物质进行输送,所以细胞壁的结构具有重大意义。除此之外,细胞壁还含有蛋白质、脂肪、矿物质。

- 细胞膜
- 紧贴细胞壁的内面,厚度约150nm,是一层半透性的膜,构成细胞壁的基础物质。细胞膜调节着细胞内的渗透压,调节着营养物质的吸收和代谢物的排出,形成酵母细胞的渗透框架。同时,细胞膜可分离出胞外酶,胞外酶由酵母细胞形成,但在酵母细胞外起作用。
- 细胞膜中最重要的基础物质是磷脂,磷脂具有对其他作用而言非常重要的典型结构。每两个脂肪酸残基与甘油产生酯化反应,在甘油的第三个OH-根上,通过磷基结合了一个氨基酸(磷脂)
- 脂肪是细胞膜的主要组成物质,其形成过程消耗很多能量,同时需要氧气存在,在此过程中脂肪酸的一部分转化为熔点较低、运动性更好的不饱和脂肪酸,缺乏氧气时细胞的形成会提前停止。
- 细胞质
- 占细胞体积50%以上,是细胞内部最重要的部分,是细胞的反应中心,大多数营养物质的分解代谢及细胞自身基础物质的合成过程都在这里完成,整个中间物质代谢:糖解、脂肪酸的分解、蛋白质的生物合成以及其他过程都在这里同时进行。
- 营养物质丰富的时候,如发酵开始时,酵母细胞的储存物质会增加,糖原这种储备碳水化合物的含量可超过酵母干物质的30%,储存在细胞质中,海藻糖这种双糖也会储备,此外还有酵母形成新的细胞物质所必需的磷和脂肪。
- 细胞中通常会充满酸性细胞液和液泡,这里储存着特定的蛋白质和多余的盐类,部分为结晶形式,当外部的渗透压由于较高的浸出物或酒精含量上升时,细胞可以通过结晶盐的可逆変化调整其内部压力。
- 细胞核
- 直径为0.5~1.5μm,经染色后可以观察到。细胞核被核膜包围,其主要化学组成是脱氧核糖核酸DNA和蛋白质,是遗传物质的承载体,控制着酵母的新陈代谢。
- 液泡
- 在显微镜下,常可看见酵母细胞中充满水性细胞液的液泡,酵母细胞可在液泡中段时间储存代谢产物,此外液泡中海油细胞的磷酸盐储仓。
- 颗粒
- 细胞中的颗粒是酵母的贮藏物质和细胞的代谢产物,包括异染颗粒、肝糖和脂肪粒等。异染颗粒中含有较多的核酸或核酸化合物,主要为核糖核酸。幼细胞生活力强,不易积累,含异染颗粒较少,老细胞中积累较多。肝糖是酵母的贮藏物质,在旺盛繁殖的幼细胞中很明显。一般酵母培养48小时,肝糖含量达到高峰,用碘化钾溶液可染成棕色。脂肪粒分散于细胞中,大小不等。当酵母形成子囊孢子时,脂肪粒用苏丹溶液可染成红色。
- 线粒体
- 线粒体一般是看不到的,形状随培养条件改变而异。在好氧条件下,特别是葡萄糖含量很低时,线粒体均匀分布在细胞质内;在厌氧条件下,线粒体黏附成厚束,分布在细胞外围。线粒体含有细胞色素和呼吸酶,负责糖类的氧化代谢,分解为二氧化碳和水,同时在产生、积累和分配细胞的能量方面起重大方式
酵母的繁殖与生长
酵母的繁殖方式可分为有性繁殖和无性繁殖,其中以无性繁殖为主。
- 无性繁殖
- 芽殖
- 这是酵母菌最普遍的繁殖方式,开始由母细胞生成芽细胞,当芽细胞成长至差不多大小后从母细胞分离,形成新的细胞。


- 假菌丝
- 当处于生长旺盛阶段时,由于新生成的芽细胞还没有来得及从母细胞中分离便又生成新的芽细胞,这样便形成了酵母菌的假菌丝。


- 裂殖
- 裂殖是动物细胞生殖的最常见的方式,也是酵母菌的一种生殖方式,首先母细胞核分裂为两个,然后这两个细胞核向两边拉伸母细胞,最终分裂为两个新的细胞。


- 有性繁殖
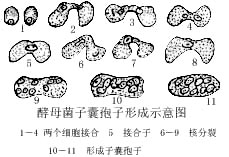
- 两个酵母菌通过接触并融合后形成子囊孢子,子囊孢子在十一的环境下萌发,然后孢子分离母细胞后开始生长,并重新以出芽的方式进行繁殖。

- 酵母的生长
- 酵母菌在生长繁殖过程中也遵循幼而壮,壮而老,老而衰,衰而死的生物生长规律。把一定数量的酵母菌,接种于合适的液体培养基中,在适宜的温度下培养时,它的生长过程具有一定的规律性。在其生长过程中,定时取样计算酵母细胞数量,然后以酵母细胞数的对数作纵坐标,生长时间作横坐标,绘制所得的曲线图称为生长曲线。


- 分析酵母的生长曲线,大致可划分为四个阶段:
- 迟缓期
- 酵母菌刚接入新的培养液中,一般并不立即开始生长,需要经过一定的时间才开始生长,这是由于细胞内各种酶系要有一个适应的过程,这段时间就叫做迟缓期或者调整期。迟缓期的长短与接入菌种的老幼、培养液的成分、菌种的种类有关。如果菌种来自生长旺盛的培养液,麦芽汁温度适宜,适当充气供氧,麦汁营养充足,则迟缓期短,反之则长。在此期间,酵母细胞并不增加(即增值速度为零),但每个细胞体积增大,细胞质变得更加均匀,贮藏颗粒物质逐渐消失,代谢机能变得非常活跃,然后开始出芽繁殖,生长速度逐渐加快,时间可能需要数小时。
- 在生产中,为缩短生长周期,通常希望调整期时间短一些,可以通过增加接种数量、采用生理态处于对数生长期的酵母细胞做种子等措施来实现。
- 这里可以添加一个“加速期”,此阶段紧接着迟缓期,细胞分离速度加快
- 对数生长期
- 酵母经过迟缓期后,就以最快的速度进行繁殖,进入对数生长期。此时,酵母每增殖一次,所需时间最短,酵母在恒定时间内,每个细胞进行芽殖,由一个细胞变两个,两个细胞变四个,即以2n的速度增长,指数n代表酵母细胞增殖世代(细胞生殖的一个周期)的数目。无论开始时细胞数目多少,其繁殖率总是2n。新细胞增殖所需要的时间称为世代时间(G),繁殖的每个世代的时间随菌种和培养条件相差甚大。
- 如果酵母培养的总时间为t,以Xo表示接入培养基的酵母数,经过t时间培养后,测得其细胞总数为Xt。公式:Xt = Xo × 2n
- 由此可求出n,即在t时间内增殖世代数:
- lgXt = lgXo + nlg2
- n = (lgXt - lgXo) / lg2
- = 3.32lg(lgXt / lgXo)
- 因为世代时间 G = t / n,由此可求:
- G = t / (3.32lg(Xt / Xo))
- 由此可求出n,即在t时间内增殖世代数:
- 在对数生长期中,酵母细胞数目的增加与时间成正比关系。在对数生长阶段,世代时间是稳定的。酵母在最适条件下生长的世代时间为2小时。影响世代时间的主要因素除菌种本身外,营养物质的浓度是主要的,此外还有培养温度、ph、渗透压和代谢产物浓度等因素。处于对数生长期的酵母细胞,其个体形态和生理特性比较正常,代谢旺盛,生长速度恒定。
- 这里可以添加一个“稳定期”,此阶段紧接着对数生长期,由于各种因素,比如底物减少,抑制生长的代谢物增加等,此阶段酵母细胞的增值速度逐渐减小
- 稳定期
- 酵母菌体培养到一定时间大量繁殖后,培养基内的营养物质逐渐耗尽,代谢产物(如酒精、二氧化碳等)积累,ph也改变,使之对细胞快速增值产生抑制作用,细胞不再以指数率无限地生长,自溶数和繁殖数两者达到平衡,维持一个动态平衡,使酵母细胞数维持为恒定值。
- 衰亡期
- 酵母菌生长到后期,培养基中养分大量消耗,代谢产物更多,环境条件对酵母生长逐渐不利。此时酵母细胞大量衰老,细胞自溶不断增多,活酵母逐渐减少。末期,繁殖几乎等于零,死亡率急剧增加。
- 生产中为了缩短发酵时间,往往采取各种措施使酵母及早进入对数生长期。如在酵母扩培阶段直接扩培至生产规模,选择在酵母增殖的对数生长期进行接种,以缩短酵母增殖的调整期。
酵母所需的营养
酵母细胞由多种化学物质组成。其中最主要的是蛋白质、核酸、多糖和脂类这四类大分子,他们约占细胞干重的96%。其余就是组成它们的单体以及无机盐等。水约占70%,此外还有有机酸、维生素、激素等有机化合物。这些化学物质均由碳、氢、氧、氮、磷、硫以及其他为数不多的化学元素构成。这些化学元素都来自于胞外环境。酵母细胞利用含这些化学元素的物质制造其细胞物质和组分,并进一步将它们组织成细胞结构。 在微生物的营养中有以下6大要素物质,它们是:碳源、氮源、能源、生长因子、无机盐、水。
- 碳源
- 碳是微生物细胞需要量最大的元素。能提供微生物营养所需碳元素的营养物质称之为碳源。能被微生物用作碳源的物质种类极其广泛,简单的无机含碳化合物(CO2、NaHCO3和CaCO3等)、比较复杂的有机物(烃类、醇类、羧酸、脂肪酸、糖及其衍生物、杂环化合物、氨基酸和核苷酸等)、复杂的有机大分子(蛋白质、脂类和核酸等),乃至复杂的天然含碳物质(牛肉膏、蛋白胨、花生饼粉、糖蜜、石油及其不同的馏分等)都可以被不同的微生物利用。甚至像二甲苯、酚等有毒物质均可以被少数微生物用作碳源。不同营养类型微生物利用不同的碳源。为数众多的异养微生物常利用某类有机化合物中的一种或几种物质作为它们的碳源。
- 糖类是微生物利用最广泛的碳源,尤其是葡萄糖。其次是醇类、有机酸和脂肪酸等。
- 氮源
- 氮是微生物细胞需要量仅次于碳的元素。能提供微生物生长代谢所需氮元素的营养物质称为氮源。能被微生物用作氮源的物质种类也很广泛:有分子态氮、氨、铵盐和硝酸盐等无机含氮化合物;尿素、氨基酸、嘌呤和嘧啶等有机含氮化合物。
- 无机盐
- 微生物还需要很多其他元素,如P、S、K、Mg、Ca、Na和Fe等大量元素,以及Co、Zn、Mo、Cu、Mn、Ni和W等微量元素。这些元素大多是以无机盐的形式提供的。其中,Mg、K、Na、Fe、Co、Zn、Mo、Cu、Mn和Ni等金属元素来源于无机盐的阳离子,而P、S等非金属元素绝大多数来自于无机盐的酸根。
- 无机盐的需要量虽然远小于C、N,但其重要性并不亚于它们。它们的生理功能可归纳为:提供微生物细胞化学组成中的重要元素,如P和S分别为核酸与含硫氨基酸的重要组成元素;参与并稳定微生物细胞的结构,如P参与的磷脂双分子层构成了细胞膜的基本结构,Ca参与细菌芽孢结构的皮层组成,Mg有稳定核糖体和细胞膜的作用;与酶的组成和活力有关。如Fe是细胞色素氧化酶的必要组分,Mg、Cu和Zn等是许多酶的激活剂;调节和维持微生物生长过程中诸如渗透压、氢离子浓度和氧化还原电位等生长条件,如Na和K有调节细胞渗透压的作用,由磷酸盐组成的缓冲剂能保持微生物生长过程中ph值的稳定,含S的Na2S和含疏基的疏基乙酸、半胱氨酸、谷胱甘肽和二硫苏糖醇等可降低氧化还原电位;用作某些化能自养型细菌的能源物质;用作呼吸链末端的氢受体。
- 生长因子
- 许多微生物除了上述的物质以及能源之外,还必须在培养基中补充微量的有机营养物质才能生长或者生长良好,这些微生物生长所不可缺少的微量有机物质就是生长因子。生长因子有维生素、氨基酸、嘌呤碱和嘧啶碱、卟啉及其衍生物、固醇、胺类、C2~C6直链或分支脂肪酸等。一些特殊的辅酶也能用作生长因子。能提供生长因子的天然物质有酵母膏、蛋白胨、麦芽汁、玉米浆等等。
- 生长因子的主要功能是提供微生物细胞重要化学物质(蛋白质、核酸和脂质)、辅助因子(辅酶和辅基)的组分和参与代谢。
- 水
- 水在细胞中的存在形式分为结合水与自由水:结合水是细胞结构的重要组成成分,用以保持生物活性(种子细胞中的结合水如果失去,那么将不再可发芽);自由水如下所述(失去自由水只要再次吸水后仍可在发芽)。
- 水是微生物营养中不可缺少的一种物质。这并不是由于水本身是营养物质,而是应为水是微生物细胞的主要化学组成:水是良好的溶剂;水具有运输物质的作用;水本身参与许多化学反应;水是良好的热导体,保证细胞内的温度不会因为代谢过程中能量的释放而骤然上升;水还有利于生物大分子结构的稳定;
酵母的繁殖和生长可划分为六个不同阶段。
- 调整期
- 也称为起始阶段,是进行新陈代谢的活化过程。此阶段时间的长短波动很大,主要取决于有机体类型、培养代数、培养条件等因素。细胞一旦开始分离就标志着此阶段的结束。
- 加速期
- 此阶段紧接着调整期,细胞分离速度较快。
- 对数增长期
- 在此阶段,细胞呈对数增殖,最大增殖速度保持恒定。此时形成新的一代所需时间短(即细胞数翻倍的时间)。在最佳增殖条件下世代时间为90~120分钟。
- 减速期
- 由于因素,如底物减少,抑制生长的代谢物增加等,对数增长阶段有一定的时间限制,随后进入增殖速度减小的减速期。
- 稳定期
- 这一阶段微生物的数量保持恒定。形成的信息报数与死亡的细胞数相等。
- 死亡期
- 在此阶段,细胞死亡数多于形成的新细胞数,细胞数减少。
在以上六个阶段中,每个生长阶段的时间长短和强度主要受底物、温度和酵母生理状态的影响。底物必须含有生长必需的营养物。同样,底物的水分含量、ph和氧气浓度对生长也很重要。
酵母的新陈代谢
生命的典型特征是生长和繁殖,维持生命需要持续的物质转化,即新陈代谢。其作用有以下两个方面:吸收可利用的物质作为营养,将其转化为机体本身的物质;获得生命功能所需要的能量。 为保证新陈代谢功能的进行,酵母必需有机物质,特别是糖形式的碳水化合物。酵母既可以在有氧的情况下利用糖(耗氧性),又可以在无氧的情况下分解糖(厌氧性)。耗氧且释放能量多的过程称为呼吸,厌氧且释放能量的过程称为发酵。通过呼吸和发酵获取能量的反应过程非常复杂且步骤繁多,每个反应步骤都由特殊的酶催化。在酵母细胞中,酶以一定的细胞结构连接。酶的呼吸链主要在线粒体上,而酶的发酵主要在细胞质的基础物质中进行。有机物的呼吸或发酵是以细胞内容物的输送为前提条件的。酵母细胞通过细胞壁吸收营养物质,由细胞膜进行调节。酵母细胞只能吸收与输送机理相适应的物质,而这又取决于酵母细胞中酶的多样性。
- 碳水化合物的代谢
- 在碳水化合物中,只有糖分能提供酵母呼吸或发酵。区别各种酵母的重要标准是它对不同糖分的呼吸或发酵能力。原则上所有能被酵母发酵的糖,也可以被酵母呼吸消耗;反之则不行。酵母对糖进行耗氧分解还是厌氧分解,这主要取决于有无氧气存在,在有氧情况下,酵母通过呼吸能取得能量;而在无氧情况下,则进行发酵。酵母是唯一能从呼吸转变到发酵的生物,正是基于这种转变才有了千百年的酒精饮料生产。
- 快速起发对酵母能量消耗很大,所以在发酵开始前必须给酵母提供足够的氧气,以使酵母获取能量进行发酵。而在后面的发酵及成熟阶段,生产过程在无氧状态下进行。对于啤酒酵母来说,主要碳水化合物的来源是低分子糖。酵母可以利用许多单糖、双糖和寡糖。而聚糖如淀粉和纤维素,则不能被酵母利用。了解哪些糖能被酵母发酵,这对啤酒酿造来说十分重要。可发酵的碳水化合物(按照酵母利用的顺序)如下:
- 单糖:葡萄糖、果糖、甘露糖、半乳糖。
- 双糖:麦芽糖、蔗糖。
- 三糖:棉籽糖、麦芽三糖(并非所有的酵母都能利用)。
- 一小部分糖没有被发酵,而是以化学能量的形式储存于酵母细胞中,必要时用于维持生命功能。细胞中最重要的化学储存物是腺苷二磷酸(ADP)和腺苷三磷酸(ATP),ATP参与每个生命过程,是生命所必需的能量储存物和转载物,没有ATP,酒精发酵根本不可能进行。
- 蛋白质的代谢
- 酵母需要氮化合物来合成酵母细胞自身的蛋白质。在无机氮中,酵母主要利用铵盐,但麦汁中的铵盐含量很少,酵母的主要氮源为氨基酸和低分子肽。
- 酵母不能直接将麦汁中的氨基酸合称为自身细胞蛋白质。蛋白质代谢过程由一系列复杂的生化过程组成。因此这些转化过程与发酵副产物的形成密切相关,如高级醇、连二酮、酯和有机酸等。由氨基酸形成高级醇即所谓的杂醇油就是这种转变的一个实例。氨基酸脱羧形成高级醇,亮氨酸脱羧可形成异戊醇。
- 酵母新陈代谢产物的形成以及分解取决于许多因素,如温度、压力、ph等。发酵副产物的含量对啤酒的口味及风味影响很大。
- 矿物质的新陈代谢和生长因子
- 酵母的新陈代谢还取决于足够的矿物质和生长因子,这些物质的作用不可低估。部分离子对酶促反应影响很大,如钾离子与ATP仪器促进所有的酶促反应,对于能量代谢和细胞壁的物质输送很重要;钠离子使酶活化,在细胞膜的物质输送中起重要作用;钙离子可以被锰离子、镁离子所取代,延缓酵母退化,促进凝固物的形成;镁离子对有磷参与的反应十分重要,特别是发酵中不可取代;钙离子很少量就会抑制某些酶;铁离子对酶的呼吸代谢很重要,可促进酵母出芽繁殖;锰离子在代谢中可取代铁离子,可促进细胞繁殖和细胞形成;锌离子有利于蛋白质的合成,其需求量为0.2mg/L麦汁,缺锌可使发酵出现问题;硝酸根离子可被细菌还原为亚硝酸根,对细胞有毒性,极不利于发酵。
影响酵母生长的因素
微生物的生长受到它们所处环境因素的影响极大。微生物可能在某些有害条件下不能声张,但却可以忍受而不至于死亡,因而必须区分环境条件对微生物存货的影响与对微生物生长、分化和繁殖的影响之间的差别。比如某些条件下只会降低微生物的活性,但是达到临界点后完全永久性失活。
- 温度
- 温度是影响微生物生长的一个重要因子。温度太低,可使原生质膜处于凝固状态,不能正常地进行营养物质的运输或质子梯度,因而生长不能进行。当温度升高到适宜温度时,由于细胞内酶的催化作用使得化学反应以较快的速度进行,从而生长速率加快。
- 然而,当超过临街温度时,蛋白质、核酸和细胞其他成分就会发生不可逆的变性作用。因此,当温度在给定范围内升温时,代谢和生长就会加速,当超过临界点时完全失活。每种微生物都有3中基本温度,即最低生长温度、最适生长温度和最高生长温度。根据微生物的生长温度范围,可将其分为嗜冷微生物、嗜温微生物、嗜热微生物和嗜高热微生物。
- 由于酵母菌种的不同,每种酵母都有自己独立的最适温度,一般而言上面酵母(ALE)的最适温度要高于下面酵母(LAGER),具体的酵母温度应当参考所购酵母厂商给出的指标进行控制。
- ph值
- ph影响微生物的生长。因为它影响环境中营养物质的可给态和有毒物质的毒性;影响菌体细胞膜的带电荷性质、膜的稳定性以及膜对物质的吸收能力;使菌体表面蛋白变性或水解;酶的活性;每种生物都有一个可生长的ph值范围,以及最适生长ph值。大多数自然环境ph为5~9,适合于多数微生物的生长。
- 一般而言,啤酒酵母厂商没有给出最适ph。啤酒的ph在糖化阶段一般控制在5.2左右,即使到发酵后期ph下降后,各类型的酵母均能很好的生存并工作。所以只要控制好酵母投放时麦汁的ph值,不需要过于担心ph对酵母带来的影响。
- 氧
- 微生物对氧的需要和耐受能力在不同的类群中变化很大,依据它们和氧的关系可分为几种类群。
- 好氧微生物
- 包括所有需要氧才能生长的微生物。有两类:一类是专性好氧微生物,它们的生长必须要有氧,快速分裂的细胞比缓慢分裂的细胞需要的氧更多,通常生长在培养基表面附近;另一类是微好氧微生物,它们在有少量自由氧存在条件下生长最好,因而生长在培养基表面之下的某一区域,该区域氧浓度正好符合它们生长的需要。
- 厌氧微生物
- 那些缺乏呼吸系统而不能利用氧气作为末端电子受体的微生物称之为厌氧微生物。可分为两类:耐氧厌氧微生物和严格厌氧微生物。前者是指那些尽管不需要氧,但可耐受氧,并在氧存在条件下仍能生长的类群;而后者则是指那些对氧敏感,在有氧时既被杀死的类群,所以专性厌氧微生物只能生长在氧气几乎不能达到的培养基底部附近,严格厌氧微生物并不是被气态的氧所杀死,而是由于不能解除某些氧代谢产物的毒性而死亡。
- 兼性好氧微生物
- 在有氧存在下通常进行有氧呼吸,产生CO2和水以及大量的能量,但在氧缺乏时可以转变为无氧呼吸,产生酒精、乳酸等代谢物和CO2以及少量的能量。这类微生物在有氧呼吸条件下的生长必无氧条件下的生长更旺盛,因而可以看到菌体在整个培养基中都有分布。
- 酵母菌属于兼性好氧微生物,在有氧条件下进行快速的生长繁殖并产生出大量的热和少量的CO2,在无氧条件下产生酒精和CO2。
啤酒酵母和啤酒质量的关系
在啤酒生产中酵母菌体不是最终产品,但对最终产品的质量非常重要,发酵过程中一系列的复杂生化反应均系酵母营养代谢作用而致,故酵母对啤酒生产和发酵质量,乃至啤酒的理化性能和其风味典型性,均有重要影响。
酵母与发酵速度
酵母的发酵速度是啤酒生产的重要指标之一。在一定的工艺技术条件下,酵母对麦汁顺利地完成发酵,为啤酒的质量提供了保证。同时,在保证发酵质量的前提下,充分发挥酵母细胞内在的潜力,可加速生产周期的循环,提高生产能力。 影响酵母发酵速度的因素主要有以下几个方面
- 酵母浓度
- 在充分搅拌并使酵母细胞均匀分布于发酵液中的情况下,发酵速度与酵母的浓度成正比例关系,如:
- 接种量0.6L/hl,发酵天数为9天;
- 接种量1.0L/hl,发酵天数为7天;
- 接种量2.0L/hl,发酵天数为4~5天;
- 当然,在实际生产中无需采用过高的酵母接种量,因为过高接种量容易使酵母衰退,发酵现象也不易控制。
- 酵母发酵力
- 发酵力用来衡量酵母酒精发酵的能力,因为环境的影响而有很大变化,如酵母的贮存条件、细胞表面的附着物等对发酵力均有影响。而酵母的菌龄、菌株性质的不同,其发酵力也不同。酵母代谢产物的原生质毒素,也会抑制其发酵力。当发酵生成酒精超过8.5%时,发酵也会被抑制,今儿降低酵母的发酵力。
- 麦汁组成
- 麦汁中的铜被酵母吸收累积至一定程度,将使酵母衰退而减缓发酵速度;麦汁中可被同化氮源的含量也影响发酵速度,一般情况是发酵力随含氮量的增加而提高;酵母在麦汁中发酵糖分的最适合PH值是4~6,超出范围将对发酵速度产生影响。
- 发酵温度
- 温度将直接影响酵母的繁殖和发酵力,尤其是下面酵母,较高的温度能加快发酵速度。
酵母与发酵度
啤酒酵母的发酵度反应酵母对各种糖类的发酵情况。正常的啤酒酵母能发酵葡萄糖、果糖、蔗糖、麦芽糖和麦芽三糖。酿制不同类型的啤酒,需要不同的发酵度。有的酵母具有较高的发酵度,有的酵母不可发酵麦芽三糖而使发酵度降低。
在一些非正常情况下,如果酵母的发酵度降低,一方面说明酵母有变异或污染的可能,另一方面应检查麦汁成分及发酵条件是否恰当。发酵度决定了啤酒类型和口味,一般控制啤酒发酵度:外观发酵度为65%~80%,真正发酵度为55%~70%。一般来说,发酵度低的啤酒并不醇厚,只是黏口、腻厚和甜感,其保质期也短;高发酵度的啤酒多数醇厚,具备了啤酒的“酒体”。
酵母与发酵异常现象
在啤酒的发酵过程中,常常遇到一些发酵异常现象,包括主发酵和后发酵期间的异常现象
主发酵期间的发酵异常现象
裂纹现象
在主发酵期间的起泡期和高泡期,发酵液表面布满泡沫时,发生液面泡沫开裂,泡沫慢慢变薄,而且不均匀,发酵不旺盛。发生这种现象的主要原因:一方面是洗涤酵母后,贮存室水温和室温变高,促进酵母代谢作用加强而缺氧,酵母衰老,造成发酵减退;另一方面是由于麦汁中α-氨基氮含量不足,溶解氧含量少,接种温度低,以及麦汁浑浊使酵母细胞表面吸附过多的蛋白质和酒花树脂,酵母的酶不能与糖类作用,使发酵变为迟缓,从而出现了裂纹现象。
防止和解决办法:在糖化时用乳酸或磷酸调整醪液的ph值;延长和促进蛋白质休止时间;提高麦芽汁α-氨基氮含量;提高麦芽汁接种温度和麦芽汁中的氧气含量;提高酵母使用量。
泡沸现象
泡沸发酵也称为沸腾发酵。常在主发酵后期或落泡期或下酒捞出泡沫时出现,一种现象是发酵液表面的泡盖由一角或一边推向另一边,部分页面又出现白色泡沫;另一种现象是大量的二氧化碳气泡上涌,发酵液像喷泉一样剧烈翻动,把以沉淀的酵母块带到液面。在这种现象未发生前,朱发酵现象如降糖都是正常的。
泡沸发酵是时有发生的,但发生的原因还不是十分清楚,说法很多,其中比较合理的解释是:一是酵母不纯,有产生气体的微生物,随酵母沉积到底部,产生大量的二氧化碳,积聚到沉淀的酵母中,最后把酵母层冲开,急剧上升,形成沸腾;二是麦汁组成成分不良,麦汁浑浊不清,固体随酵母沉淀到底部,酵母继续利用固体的营养物质发酵产生二氧化碳,积聚在其中,量大时把酵母冲开,二氧化碳气体随之上升而沸腾;再一种是主发酵温度过高,后期采取急剧降温,当把泡盖去掉后,表面压力降低,使下边二氧化碳急剧上升造成沸腾;还有一种是啤酒酵母变异,凝聚性不良,而且麦汁可发酵性糖比例偏高,产生旺盛发酵,二氧化碳产生量多,在泡盖捞出后,二氧化碳急剧上升所造成。
对于沸腾发酵,先在技术管理上还没有有效的办法解决,一般采取改进糖化操作,改善麦汁组成成分,加强麦汁过滤,使麦汁清亮透明,另外,从加强酵母管理着手,遇到泡腾发酵时,重新培养酵母,并加强卫生管理和灭菌工作。
气泡发酵现象
气泡发酵也称为异泡发酵,即在主发酵期的低泡期,或发酵终了前一天,发酵液表面已形成的棕色泡盖上出现多数的大气泡,或称为大明泡,继而破坏了泡盖,使已凝结出来的酒花树脂和蛋白质凝结物下沉,使表面的泡盖变成白色的现象。产生气泡发酵的主要原因是酵母无染杂菌、野生酵母或其他原因,其次是糖化不完全,糖化用水中亚硝酸盐过量。出现这种情况后发酵液不易澄清,啤酒的口味不纯。
防止的办法:加强酵母室和发酵室的卫生管理工作,及时做好清洁、灭菌工作和现场使用的酵母的洗涤保管工作,及时培养新鲜强装的酵母,按期更换,加强糖化管理,对糖化用水定期检测,受亚硝酸盐和硝酸盐污染的水要进行处理,根据原料情况调整糖化操作,使生成的麦汁符合要求。
虚泡现象
在主发酵落泡期,形成疏松的泡沫,开始是白色,逐渐变成棕黄色,最后泡盖松散无力,凝结在下面的酒花树脂沉入发酵液内。这主要是由于原料麦芽溶解不好,糖化时蛋白质分解的温度和时间不恰当所造成。
防止的办法:对原料事先做好监测工作,根据原料情况制定糖化工艺操作方法,加强半成品的分析。
发酵中止现象
主发酵达到高泡期,泡沫升起不久,很快又回缩,糖度下降缓慢,甚至出现发酵中止现象,同时发酵液澄清。产生发酵中止现象的原因是多方面的,如糖化所得的麦汁α-氨基氮或微量物质嘌呤、嘧啶含量不足,可发酵性糖含量过低,糊精等非糖含量过高;糖化时醪液的PH值过高,使植酸钙和镁盐不能充分分解为肌醇和磷酸盐,使麦汁中缺乏生长素;麦汁中的酸度过高或过低,极易造成酵母沉淀;在主发酵时,温度掌握不当,突然降低温度,使酵母过度受刺激而沉淀;酵母发生变异,不能发酵麦芽三糖等。发酵中止可造成发酵液发酵度低、残糖高、有甜味、口味淡薄、不爽快、泡持性不好。
解决的办法:好次原料搭配使用,防止糖化麦汁质量不一,同时在糖化时,对糖化用水加强处理,调整水的PH值,有利于提高可发酵性糖和可同化氮化物的含量,以及生长素的含量,在糖化时促进蛋白质的分解和麦汁煮沸时的凝固;严格管理发酵工艺,防止温度忽高忽低,避免酵母受刺激。发生这种现象后可采取倒桶、添加酵母、通风搅拌、供应充足的氧气,使之重新发酵;也可将中止发酵的发酵液倒入两个发酵罐中,分别加满已繁殖后的发酵液,使之重新发酵。
再发酵现象
在主发酵末期,泡盖已经形成,忽然又开始旺盛发酵,形成白色泡沫,将已形成的棕色泡盖翻入发酵液中。发生这种现象的主要原因是,麦汁组成成分不当,可发酵性糖少,但糊精的中间产物通过酵母中酶的作用,又被酵母所利用,另一种原因是酵母变异了。
解决的办法是对所使用的酵母要做性质检查,确定酵母是否变性,目前有些厂所使用的酵母经过长期的高温发酵,降糖速度很快,一天可以降糖4~5°P,这也是酵母变性问题,另一种解决办法是调整糖化工艺。
后发酵期间的发酵异常现象
发酵不旺盛现象
发酵不旺盛,开口发酵时造成泡沫不溢出的原因主要有:下面发酵液中酵母细胞少;发酵罐中所留空隙太大;酵母衰老,发酵作用已极为微弱。解决措施:可采用添加高泡酒的方法。
贮酒罐不升压现象
贮酒罐封罐后,在3~10天内罐压应升至0.05~0.08MPa,若封罐后不升压应检查原因,如贮酒罐是否漏气;酒液中酵母细胞数是否过少;一般可采取倒罐、加高泡酒的方法进行解决。
发酵沉淀不清现象
贮酒较长时间后,酒液浑浊不清,其原因:糖化不完全,蛋白质分解欠佳;麦汁中α-氨基氮过少;PH值不当;酵母被杂菌污染。
解决措施:应重新调整糖化工艺,调整麦汁组成,或倒罐至其他新发酵的酒液罐内;若酵母感染杂菌,则应该处理掉。
酵母的检查与鉴定
啤酒酵母的质量直接关系到啤酒发酵和啤酒的质量。如果啤酒酵母被杂菌污染或发生变异,就会产生不正常的发酵现象和影响啤酒的口味。酵母的自然变异是比较低的,但是在长期的酵母培养和发酵过程中变异的可能还是存在的。
啤酒生产中酵母的检查
啤酒生产过程中,经常对酵母进行镜检和做某些生理特性试验,镜检一般只起辅助作用,对酵母某些生理特性的检查更具有重要性。
外观和形态检查
菌落形态
- 液体培养基中观察菌落形态
- 在液体培养基中观察发酵液浑浊的快慢,澄清的程度及酵母沉淀的情况。酿造车间现场使用的酵母泥必须新鲜,呈黄白色,有果实的爽快香味,其上部洗涤水透明且无色。沉于底部的酵母泥紧密,取起后应松散而不黏连。如色泽深暗或发粘则说明质量较差。
- 啤酒酵母个体形态观察
- 用显微镜观察酵母细胞的形状、大小、夹杂物以及是否有细菌等。啤酒酵母呈球形、椭圆形或卵圆形,细胞的平均直径为4~5μm,大小为(3~7)μm×(5~10)μm。液体培养的酵母细胞大于固体培养的细胞。成熟细胞大,年幼时细胞小。
- 优良健壮的酵母细胞具有均匀的形态和大小,平滑而薄的细胞膜,细胞质透明均一,年幼少壮的酵母细胞内部充满细胞质;老熟的细胞出现空泡,内贮细胞液,呈灰色,折光性强;衰老的酵母死亡率高,可通过美蓝染色,检查酵母死亡率。一般生产上使用的酵母死亡率应在3%以下,新培养的酵母死亡率在1%以下。
- 下面发酵啤酒酵母一般以单端出芽繁殖,很少形成短链。芽在脱离母细胞前总是比母细胞小,芽和母细胞的纵轴有30°夹角,这和许多野生酵母的芽和母细胞在一个纵轴上形成鱼鳔形有明显区别。在镜检中,如果发现显著变异,可怀疑是酵母退化或可能有杂菌污染,需要另做较细致的细菌或野生酵母鉴定。夹杂物为蛋白质和酒花树脂等,如与细菌等分辨不清时,可在酵母中加入10%碱溶液或50%醋酸,则蛋白质小颗粒溶解,标本中只看到酵母和细菌。
巨大菌落的观察
某些酵母的品种鉴别上,在一般的形态上不易区别。但是,它们所生成的巨大菌落则不一样,菌落越大形态越容易区别。一般,巨大菌落表面平滑多为分散型酵母,而表面褶皱都为芽簇型酵母。巨大菌落和发酵性能之间没有什么联系。有的菌种巨大菌落为白色,表面平滑呈扁平或半透明镜状隆起,有时中部略呈凹形。
杀菌及其污染程度的观察和检查
确定50个显微镜视野中存在的杂菌数并按等级确定污染程度:
| 杂菌数 | 污染等级 |
| 1个 | 微量 |
| 3个 | 很少 |
| 6个 | 少 |
| 8个 | 轻度污染 |
| >8个 | 强度污染 |
对单一微生物允许污染标准如下:
| 野生酵母 | 很少感染(3个) |
| 细菌(杆菌、乳酸菌、四链球菌) | 少感染(6个) |
| 无损害的微生物(球菌和小酵母) | 中等感染 |
按照以上标准,在每次酿造前应该精确检查,并确定污染程度和新细胞的比率。应该保持一个视野中约有100~200个酵母细胞,因此悬浊液是较浓的。
死细胞的检查
良好的现场使用酵母其死细胞数一般在0.5%~3%之间,不应超过5%。检查时取磷酸缓冲液(0.2mol/L磷酸氢二钠0.25ml和0.2mol/L磷酸二氢钾99.75ml混合)和0.04%美蓝溶液等量混合,既得0.02%美蓝染色液,其ph为4.6,此溶液需保存于暗处。同时取适量浓度的酵母悬浊液1ml(泥状酵母1ml用水稀释成200倍),混合1ml的染色液,5min内在显微镜下检查,数出被染色(蓝色)的酵母死细胞数,计算其百分率,即为酵母细胞死亡率。
肝糖染色检查
发酵力强的酵母细胞始终含有肝糖。肝糖在强盛的发酵阶段形成,其数量决定于麦芽汁的组成。如酵母长期在水下保存(3~5天)则肝糖完全消失,因为肝糖作为贮存物质一部分被酵母所消耗,同时被酶催化分解后的一部分也转移到水中。不旺盛的发酵或者发酵开始迟缓都是肝糖成分不足的标志。一般肝糖都是在纯培养时进行测定,该测定项目也可以在生产中发现发酵不良时进行。但是目前还不能精确地指出一个发酵力强的酵母应该含有染成怎样程度的褐色细胞。肝糖含量首先决定于酵母的生理形态,其次取决于生长的强度和贮存时间。发酵后取出的健康教母应有70%为染成深褐色的细胞。
异染颗粒的检查
新鲜强装的酵母异染颗粒粒大且色深,根据其含量可以判断酵母在生理学上的能力。很多学者认为,异染颗粒的含量与酵母的发酵能力和繁殖之间存在着一定的关系。异染颗粒是根据基质中磷酸化合物的存在而存在,在判断老的和退化的酵母时,对其进行检查是可行的。
芽蔟的检查
将现场使用的酵母稀释并制成悬滴镜片,在显微镜下观察酵母的连结是由于凝聚力还是由芽蔟形成所致。如果识别有困难,可在酵母中加稀薄的氢氧化钾溶液或稀醋酸溶液,然后再进行显微镜检查。此时,凝集酵母细胞即个个分离而芽蔟细胞仍在。下面发酵现场使用的酵母如果有芽蔟存在,可以人为是由于异种酵母的混入所致。
孢子形成速度的检查
将酵母移植到酒石酸蔗糖溶液中(酒石酸4g,蔗糖10g,溶于100ml水中),20℃培养48h,如此进行2~3次,再在麦汁液体培养基中繁殖24h后移植到石膏块上(石膏块应先刮平,上有凹部,放入双重皿中杀菌,以水浸没石膏块1/2处),在25℃恒温箱中放72h,用显微镜检查孢子形成情况。如是野生酵母,此时已经形成孢子。
酵母生理特性试验
发酵力
酵母发酵力用以判别其酒精发酵的能力,一般来说应该选择发酵能力强的酵母。如果酵母发酵力衰退则意味着酵母发生退化、变异。发酵力测定的方法有以下两种。
发酵法
取灭菌的250ml三角瓶,加上棉塞,瓶内加入150ml麦汁,经过常压灭菌,冷却后加入酵母泥1g摇匀,放在25℃保温箱中进行发酵,每隔8h震动1次,经过3~4天发酵终了,过滤掉酵母,取发酵液100g,并把酒精蒸馏出去,放在定量瓶中添加蒸馏水使重量恢复到100g,混合摇匀,测量20℃时的密度,测出残留在发酵液中的浸出物浓度,利用下面的公式计算真正发酵度:
- 真正发酵度(%)=(发酵前麦汁浓度 - 发酵后排除酒精的麦汁浓度) / 发酵前麦汁浓度
发酵后,不用蒸馏法去除酒精,直接测量密度,算出发酵液残留浸出物的浓度,利用下面的公式计算其外观发酵度:
- 外观发酵度(%)=(发酵前麦汁浓度 - 发酵后麦汁浓度) / 发酵前麦汁浓度
外观发酵度一般比真正发酵度高约10%,换算方法如下:真正发酵度 = 外观发酵度×0.819。啤酒的发酵度一般都分为高、中、低三个类别,见下表:
| 淡色啤酒 | 浓色啤酒 | |||
| 外观发酵度 % | 真正发酵度 % | 外观发酵度 % | 真正发酵度 % | |
| 低发酵度 | 60 - 70 | 48 - 56 | 50 - 58 | 41 - 47 |
| 中发酵度 | 73 - 78 | 59 - 63 | 60 - 66 | 48 - 53 |
| 高发酵度 | > 80 | > 65 | > 70 | > 56 |
二氧化碳减量法
用已知重量的250ml发酵瓶装入12 ~ 18°P的麦汁150ml,在101.325Pa下灭菌15~20min,冷却后添加酵母泥1g,接上发酵栓(类似水封,里面加入杀菌液可以洗涤气体),擦干瓶外的水汽,称其重量。把发酵瓶放在20℃的保温箱中发酵,每天定时称重,发酵6~8天,最后每天减重不高于0.2g,即为发酵终止。
优良的酵母发酵力强,二氧化碳气体溢出多,一般中等发酵的酵母在12°P麦汁中发酵失重> 3.6g,若逐日比较二氧化碳失重情况,由此可判别发酵速度。
酵母热死亡温度
微生物的热死亡温度是指液态培养的微生物,在某温度下即刻被杀死,此温度称为微生物的热死亡温度。啤酒酵母一般在45℃时即停止生命活动,热死亡温度为50~54℃。
为了避免试验酵母的缺点,习惯上先将试验酵母移到液体培养基中在25℃培养24h后试用,或从扩培中采取以供实验。酵母的热死亡温度除了与培养基种类有关外,与加热时间长短也有关。啤酒厂选择的温度为40~60℃,每个间隔温度为2℃,保温时间习惯以10min为度。
若酵母的热死亡温度改变,说明菌种发生变异,或受到野生酵母污染。野生酵母比培养酵母有更高的耐热性。
酵母的凝聚性试验
啤酒酵母的凝聚性是区别菌株的一项重要内容,在生产商具有特殊的重要性。各种酵母的凝聚性有较大的差别,当酵母发生变异或衰老时,凝聚性随之发生较大的变化。啤酒酵母的凝聚性不同,酵母的沉淀速度也有差异,发酵也不一样。凝聚性强的酵母,发酵液容易澄清,但发酵度偏低;凝聚性差的酵母,发酵液不易澄清,酵母回收困难,但是发酵度高。
啤酒酵母凝聚性测定的方法采用本斯(Burns)实验法:将1g酵母泥与10ml、pH4.5的醋酸缓冲液混合,20℃平衡20分钟,加至带刻度的锥形离心管内,连续20分钟,每隔1分钟记录沉淀酵母的容量。实验后,检查pH是否保持稳定。一般规定10分钟时的沉淀酵母量在1.0ml以上者为强凝集性,0.5ml以下者为弱凝集性。
发酵速度的测定
发酵速度与酵母品种有关,如酵母的麦芽糖酶活性是控制麦芽糖发酵的重要因素,与发酵速度关系很大;发酵速度与环境条件的关系也很密切,如麦汁成分、发酵温度、通风条件、发酵容器等。一般来说酵母在统一条件下发酵速度越快越好。
为了取得与现场发酵条件相似的发酵速度,测定方法是:在直径5cm、长120cm的玻璃筒内,装2L麦汁,接种后按现场发酵条件控制,每天测定其外观浓度,观察对比其发酵速度。
感官鉴定
不同的啤酒酵母,其发酵时的代谢产物不尽相同,因而发酵液的风味也不一样。只有优良的啤酒酵母才能产出优秀风味的啤酒,不仅风味好,还要有正常的芳香,而且要求风味始终保持一致,如果生产过程中产生怪味和异味,就必须检查所用酵母是否发生变异或污染。
耐酒精浓度的试验
酵母在麦汁中发酵,到某一程度即停止。其原因,一是由于可发酵性糖的耗尽,二是受酒精含量的抑制。这在实际应用上具有很大意义。虽然在通常的啤酒发酵中,酒精含量较低,对酵母的影响不大,但不同的酵母对酒糟浓度的耐受力各有不同,在发酵时,一般采用能耐受较高酒精度的酵母,以有利于发酵。
染色实验
通过美蓝染色试验计算其死亡率。新酵母(包括或扩培后的酵母)的死亡率应当< 1%,现场使用中的酵母死亡率应当< 3%。
降糖速度
啤酒酵母的发酵能力可用降糖速度来表示。正常培养的酵母,第10天外观浓度应该不高于3.5°P,如果在4.0°P以上则为降糖慢的酵母。